PERSPECTIVES ON THE “LIVING PLESIOSAUR” CONTROVERSY
Edited by and featuring commentary by Scott Mardis
“Loch Ness Monsters (Nessies) are—if they exist—animals of a species either not yet known to science or known but thought to have been long extinct.
Much controversy has concerned eyewitness testimonies and photographs whose relevance and validity are uncertain. However, there also exists a body of objective evidence that critics have been unable to gainsay: the Dinsdale film; numerous sonar echoes obtained over many years by different investigators; and underwater photography in 1972 coincident with sonar detection of large targets.
If the descriptions of Nessies provided by photos and eyewitnesses could be interpreted as some species of animal known from anywhere else in the world, there would be no great fuss about it. If sharks, say, or dolphins, or some small whales had adapted to fresh water, that would be quite interesting to biologists but no reason for world-wide media or public interest. The trouble is, Nessies look like nothing now known to be alive anywhere. Perhaps even worse, they look rather dinosaur-like. The real animals that they resemble most closely are plesiosaurs, marine creatures that once thrived in the oceans all over the globe but that are believed to have been extinct for tens of millions of years.”- Henry H. Bauer, The Case for the Loch Ness “Monster”: The Scientific Evidence, Journal of Scientific Exploration 16(2) (2002), pp. 225–246
Top image: 1975 Academy of Applied Science Loch Ness photograph, Bottom image: Skeleton of Cryptocleidus oxoniensis plesiosaur
“I think they got frightened. Those who make their living from this, the zoologists, are not ready to believe, on the basis of one picture, that something that should have been dead 65 million years ago is still existing in some form at Loch Ness, Scotland.”- Dr. Robert Rines, NOVA: The Beast of Loch Ness, PBS Television, 1999
"The publication of Scott and Rines and the photographs in the national press indicate that there may be a plesiosaur-like reptile inhabiting Loch Ness. It is exceedingly difficult to envisage how a former tropical marine reptile could endure the cold waters and harsh environment provided by a small lake in Scotland. Since Loch Ness did not exist until some 12,000 years ago, one is faced with the problem of the survival of Nessiteras for a period of 64 million years in a world where it’s former ecological niche has been occupied by modern cetaceans and pinnipeds” – L.B. Halstead, P.D. Goriup and J.A. Middleton, Departments of Geology and Zoology, University of Reading, U.K., letter to the journal Nature 259 (1976), pp. 75-76.
“What the body temperature and thermoregulation processes of extinct vertebrates were are central questions for understanding their ecology and evolution. The thermophysiologic status of the great marine reptiles is still unknown, even though some studies have suggested that thermoregulation may have contributed to their exceptional evolutionary success as apex predators of Mesozoic aquatic ecosystems. We tested the thermal status of ichthyosaurs, plesiosaurs, and mosasaurs by comparing the oxygen isotope compositions of their tooth phosphate to those of coexisting fish. Data distribution reveals that these large marine reptiles were able to maintain a constant and high body temperature in oceanic environments ranging from tropical to cold temperate. Their estimated body temperatures, in the range from 35° T 2°C to 39° T 2°C, suggest high metabolic rates required for predation and fast swimming over large distances offshore.
Paladino et al. proposed that some marine reptiles such as leatherback turtles display endothermy instead of inertial homeothermy, thus helping them to feed in cold waters. However, Lutcavage et al. showed that the studied gravid female specimens raised their metabolic rates because of egg laying, thus biasing the evaluation of their true metabolic status.
Adaptation to cold marine waters was also revealed by the fossil reptile assemblage discovered in the Aptian southern high-latitude deposits of the White Cliffs in southeast Australia. The specimens were attributed to at least three families of plesiosaurs and at least one of ichthyosaurs. Paleoclimatic proxies indicate cold to near-freezing conditions at the seasonal scale, a climate mode that is not tolerated by modern ectothermic reptiles such as turtles or crocodiles. This observation suggests that some Mesozoic marine reptile taxa were able to cope with low temperature marine environments.”- Aurélien Bernard, et al. Regulation of Body Temperature by Some Mesozoic Marine Reptiles, Science 328(2010);pp.1379-1382
(from Ryosuke Motani, Warm-Blooded “Sea Dragons”?, Science 328, 2010, pp.1361-1362)
(Left) Mesozoic Geologic Table. (Right) South Pole (Early Cretaceous Period).
“The subsurface opal-bearing deposits of the Bulldog Shale at Coober Pedy and Andamooka in South Australia, and the Doncaster Member of the Wallumbilla Formation at White Cli
ffs in New South Wales, represent localities of some significance, producing ichthyosaurs and a high diversity of plesiosaur taxa. The sediments comprise predominantly finely laminated shaly mudstones and claystones representing deposition under transgressive shallow marine conditions, and in an Early Cretaceous high latitude zone (60(–70(S;). Because of severe weathering and often-poor locality data (a result of specimens being discovered serendipitously and extracted during opal mining), assignment of most marine reptile fossils to exact stratigraphic horizons is again problematic. However, a predominantly Aptian age can be suggested on the basis of macro invertebrate assemblages , and the presence of potentially ice-rafted quartzite/porphyritic boulders and glendonite (pseudomorphs of the calcium carbonate hexahydratemineral ikaite) nodules.
These conspicuous sedimentary structures characterise the early depositional stages of the Bulldog Shale and Wallumbilla Formation in the southern Eromanga Basin and have been correlated with a period of very cold to near-freezing climatic conditions during the Late Neocomian–Early Albian. A similar cool temperate to very cold environmental setting has been suggested for the Lower-mid Albian (C. paradoxa Zone) estuarine–coastal plain facies of the Griman Creek Formation at Lightning Ridge, New South Wales/Suratregion, Queensland, and the Aptian–Lower Albian (C. hughsii Subzone–C. striatus Subzone) braided stream and overbank flood plain deposits of the Wonthaggi and Eumeralla formations in Victoria. These units have produced a handful of plesiosaur remains (mainly isolated teeth) that represent animals living near to or within the Cretaceous southern polar circle and evidently adapted to at least seasonal occupation of inland freshwater environments.
Leptocleidus is currently the most widely distributed plesiosaur genus known from Australia. Isolated remains also possibly attributable to Leptocleidus have been described from Albian freshwater sediments of the Griman Creek Formation at Lightning Ridge in New South Wales and the Surat district of Queensland. The bulk of these specimens consist of isolated teeth, although some skeletal elements, including a large propodial similar to that of Leptocleidus clemai (from the Hauterivian–Barremian Birdrong Sandstone of Western Australia),have been discovered. Fragmentary pliosauroid teeth and ribs closely resembling those of Leptocleidus sp. have also been recorded from the freshwater braided stream and overbank floodplain deposits of the Wonthaggi and Eumeralla formations in Victoria. Interestingly, these deposits were laid down in a very cold, high-latitude environment and provide evidence of plesiosaurs living within inland streams and rivers at the Cretaceous South Pole.”- Benjamin P. Kear, Cretaceous marine reptiles of Australia: a review of taxonomy and distribution, Cretaceous Research 24 (2003), pp. 277–303The record of plesiosaurians from freshwater deposits is sparse in comparison to those from marine sediments. Despite this, a number of discoveries have been made from around the world. The fact that these range in age from Early–Middle Jurassic to Late Cretaceous attests to the group’s long history of habitation in non-marine environments.
The vast majority of non-marine plesiosaur specimens are fragmentary, and many are taxonomically uninformative. Where they are diagnostic, however, many of the freshwater specimens are referable either to ‘rhomaleosaurid-like’ taxa, or to the widespread Cretaceous pliosauroid genus Leptocleidus. Not surprisingly, therefore, the material from southeastern Australia shares similarities with this latter taxon, and lends support to the hypothesis that freshwater and near-shore marine environments may have served as refugia for plesiomorphic pliosauriform plesiosaurs well into the late Early Cretaceous. The plesiosaur fossils from southeastern Australia constitute one of a number of recognized finds from Cretaceous high-latitude deposits. However, most other occurrences are marine in origin, including examples from central Australia, New Zealand, the Chatham Islands, Patagonia, Antarctica and the Canadian Northwest Territories. Amongst the currently documented specimens, those from the Early Cretaceous units of central and southeastern Australia are unusual because they occur in association with paleoclimatic indicators (e.g., cryoturbated sediments, glacial erratics, glendonites, and growth-banded wood) denoting seasonally very cold to near freezing conditions. This contrasts markedly with climatic regimes typically tolerated by modern aquatic reptiles, but suggests that some plesiosaur taxa may have been able to cope with extremely low average water temperatures.”-Benjamin P. Kear, PLESIOSAUR REMAINS FROM CRETACEOUS HIGH-LATITUDE NON-MARINE DEPOSITS IN SOUTHEASTERN AUSTRALIA, Journal of Vertebrate Paleontology 26(1) (2006), pp.196–199

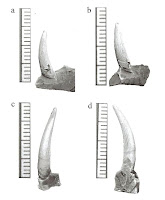
Freshwater plesiosaurs from southeastern Australia (from Kear 2006) The taxonomic status of three previously reported plesiosaurian specimens from the non-marine Horseshoe Canyon Formation (Campanian-Maastrichtian, Upper Cretaceous) is reassessed. The holotype of Leurospondylus ultimus and a previously undescribed partial skeleton from Red Deer River Valley represent indeterminate elasmosaurids. The two plesiosaurian specimens came from different horizons for which different environments are inferred. The larger individual may be closely related to Elasmosaurus. This study supports the hypothesized decline of polycotylid plesiosaurs in the North American during the Late Campanian and suggests small adults and juveniles of elasmosaurids existed in both marine and non-marine environments whereas large adults were limited to the latter.”- Tamaki Sato and Xiao-Chun Wu, REVIEW OF PLESIOSAURIANS (REPTILIA: SAUROPTERYGIA) FROM THE UPPER CRETACEOUS HORSESHOE CANYON FORMATION IN ALBERTA, CANADA, Paludicola 5(4) (2006), pp.150-169

Freshwater plesiosaurs, probably Elasmosaurid, from Horseshoe Canyon, Alberta, Canada(from Sato and Wu 2006)
“While the majority of plesiosaur material is associated with marine shelf deposits, some specimens have been found associated with lagoonal, estuarine or delta-margin environments and freshwater environments. Sedimentological evidence for the environment of deposition of the Axel Heiberg locality indicates that it was deposited in a lagoon or bay, but the evidence is equivocal as to the salinity of the water during deposition. While the occurrence of the plesiosaur suggests marine or brackish-water conditions, the associated fauna suggests a non-marine environment. Taxa found in association with plesiosaurs recovered from marine and near-shore lagoonal environments typically include a diverse assemblage of marine vertebrates. Mosasaurs (Kear, 2003) are typically found in association with plesiosaurs in Late Cretaceous vertebrate assemblages. Other marine vertebrates typically found in association with plesiosaurs include marine turtles and various fishes, such as Enchodus, Coelodus, Oseroides, and Hoplopteryx. However, such taxa are absent in the Axel Heiberg assemblage. At this locality, the vertebrates found in association with the plesiosaur remains are all taxa that are typical of non-marine vertebrate assemblages.Thus the faunal evidence suggests that these teeth represent an additional non-marine occurrence of plesiosaurs.
Plesiosaur occurrences in non-marine settings are typically dominated by small individuals. In the Late Campanian Dinosaur Park Formation of Alberta, all the plesiosaur remains recovered are from individuals of small size, and one-third of the specimens show features indicative of juvenile individuals. The small size of the plesiosaur from the Axel Heiberg assemblage is consistent with the dominance of juveniles in non-marine environments. The Cretaceous saw an explosion of plesiosaurian diversity, and distributions extended to above the Arctic Circle. In the Southern Hemisphere, high-latitude occurrences of Cretaceous plesiosaurs have been reported from Seymour Island of the Antarctica Peninsula, New Zealand, Argentina, and Australia. In addition to the new Axel Heiberg occurrence, high-latitude occurrences of plesiosaurs in the Cretaceous of the Northern Hemisphere include reports of plesiosaurs from the Kanguk Formation of Ellesmere Island and from Upper Cretaceous sediments on Banks Island, Eglinton Island and Melville Island in the western Canadian Arctic. The ages of some of the western Arctic occurrences are not yet well constrained, but it is possible that they are from sedimentary correlates of the Kanguk Formation. Seasonal migration of vertebrates has been discussed for some Cretaceous Arctic vertebrates. But the high-latitude distribution of plesiosaurs may also be a further reflection of the extreme Turonian-Coniacian climatic warmth, which is apparent from other vertebrates found in the Axel Heiberg assemblage” – Deborah Vandermark et al., Late Cretaceous Plesiosaur Teeth from Axel Heiberg Island, Nunavut, Canada, Arctic 59 (1) (2006), pp.79-82

(Left) Freshwater Elasmosaurid Teeth, Axel Heilberg Island, High Canadian Arctic (from Vandermark et al. 2006) (Right) Leatherback Turtle (Dermochelys coracea) div>
“There used to be a time when the BBC’s ability to produce good science documentaries was highly respected: they generally portrayed a balanced view of the subject, had their facts straight, and were careful not to cast ideas, or scientists themselves, in an inappropriate light. Alas, those days are long gone, as viewing of any Horizon documentary made within the last five years will show. The BBC’s Loch Ness documentary shown last Sunday was no exception, though let me say that I like Steve Leonard and I enjoy watching his stuff. When wondering if a hypothetical plesiosaur could survive in waters as cold as those in Loch Ness, Leonard pointed to leatherbacks and to Australian fossil sites yielding (apparently) coldwater plesiosaurs.
Leatherbacks really can swim in cold boreal seas, but despite initial results from Paladino et al. (1990) they are no longer thought to have a resting metabolic rate elevated compared to that of other similar-sized reptiles. In other words, they are bradymetabolic. However, they are endotherms and have a bunch of features that allow them to be cold tolerant including an insulative carapace, a thick, oil-saturated skin, fibrous fatty tissue and counter current heat exchange mechanisms in the flippers. The presence or absence of all these things can be determined from fossils and there are enough good plesiosaur fossils (including at least one good specimen with skin impressions- my info on it is from Arthur Cruikshank, an expert on plesiosaur anatomy) for us to be confident that these things were absent in plesiosaurs. The bone microtexture and histology of plesiosaurs is not consistent with endothermy- they appear to have been ‘traditional’ reptiles. So the only getout for having them as cold tolerant is to argue that they were gigantotherms.
Problem: the entire theory of gigantothermy is based on Paladino et al.’s erroneous data on leatherbacks, and it has since been retracted by these authors. Given that other studies do not show leatherbacks to have an elevated metabolic rate (Lutcavage et al. 1990), the theory of gigantothermy has died a death, and there is no evidence that giant bradymetabolic vertebrates converge in physiology with giant tachymetabolic ones. Even if gigantothermy were a viable theory, in marine reptiles it is only theoretically possible with the suite of features cited above. Indeed Orenstein (2001) writes ‘Gigantothermy…would not be enough to keep a leatherback warm in cold northern waters’ (pg. 134). At the moment the idea that plesiosaurs may have been cold tolerant is not based on any good evidence. As for the evidence showing that some Cretaceous plesiosaurs inhabited cold water with icebergs etc., this is controversial: the Cretaceous poles were nowhere near as cold as the modern ones and the evidence that the Australian sites they referred to were frequented by icebergs rests on the presence of drop stones- rocks alien to the local sedimentary geology and which appear to have been carried to their new home by ice. The problem is that icebergs are not the only way in which drop stones get dropped. Stones and rocks can also be carried for miles and miles in the roots of floating trees and as seaweed holdfasts.”- Darren Naish, CZ Conversations: Darren Naish on Plesiosaurs, Basilosaurs and the Problems with Reconstructions, North American BioFortean Review 5 (3) (2003), pp.10-19 “Higher metabolic rates for these ancient reptiles, relative to modern ones, have previously been suggested, on the basis of bone histology and swimming energetics. All three groups (plesiosaurs, ichthyosaurs and mosasaurs) had a higher body temperature than co-occurring fish by about 5° to 20°C, with the exception of Triassic ichthyosaurs. This suggests that they had heat conservation systems such as blubber layers and specialized blood circulation.”-Ryosuke Motani, Warm-Blooded “Sea Dragons”?, Science 328 (2010), pp.1361-1362
“No other known extant reptile shows this combination of chondro-osseus developmental features. However, certain extinct reptiles show some similarities to the leatherback: plesiosaurs have been described as having endochondral and periosteal cones that do not remodel; ichthyosaurs, plesiosaurs, nothosaurs and mesosaurs are known to have amedullary bones with cancellous-compacta differentiation paralleling that of marine mammals, and protostegid turtles have also been noted briefly to be somewhat similar in this respect. All these fossil reptiles were highly adapted to a marine existence as shown by other skeletal features. No terrestrial vertebrate has these chondro-osseus developmental features. The fact that such diverse groups as cetaceans, sirenians, pinnipeds, penguins, extinct marine reptiles and amphibians, and leatherback turtles have such a high degree of physical similarity in bone morphology suggests an underlying mechanism of marine adaptability which has led to a highly developed pattern of skeletal evolutionary convergence.”-Anders G. J. Rhodin et al., Chondro-osseus Morphology of Dermochelys coreacea, a marine reptile with mammalian skeletal features, Nature 290 (5803) (1981), pp. 244-246
 |
| The Plesiosaur Polycotylus latippinus giving birth |
“Among modern reptiles, the plesiosaur-like trait combination of viviparity, small brood size, and large birth size is rare, but it does occur in the scincid Egernia species group. Because both cetaceans and Egernia-group lizards are highly social and engage in substantial maternal care, plesiosaurs may have behaved similarly. We hypothesize that large plesiosaur fetus size may indicate that plesiosaurs lived in gregarious social groups and engaged in parental care.”- F. R. O’Keefe and L. M. Chiappe, Viviparity and K-Selected Life History in a Mesozoic Marine Plesiosaur (Reptilia, Sauropterygia), Science 333(2011), pp. 870-873
“The ability of plesiosaurs to move on land is another point of contention. The mechanics of their skeletons imply a completely aquatic existence: the limb girdles are only weakly connected to the axial skeleton and this would inhibit the transfer of force from limb strokes into movement on land. However, small plesiosaurs may have been relatively unaffected by these constraints and might have used their powerful limb downstrokes to propel themselves forward in short ‘hops’.”- Adam Stuart Smith, Fossils Explained 54: Plesiosaurs, Geology Today 24(2) (2008), pp.71-75
The plesiosaur Cryptocleidus oxoniensis on land from the BBC’s Walking With Dinosaurs, Copyright Dave Martill and Darren Naish 2000

http://forteanzoology.blogspot.com/2010/03/dale-drinnon-amendment-to-arthur-grants.html
“An hypothesis requires two elements. It has to be 1) verifiable - supported by evidence and 2) falsifiable - capable of being shown false by evidence. For example: "Plesiosaurs became extinct at the end of the Cretaceous period" is a useful hypothesis. It is verified by the fact that we have not found any plesiosaurs in post-Cretaceous deposits. It can be falsified by the discovery of plesiosaurs in post-cretaceous deposits. So far, no plesiosaur fossils have been found in post-cretaceous deposits, so the hypothesis stands. "Plesiosaurs did not become extinct at the end of the Cretaceous period" sounds as if it is the corollary hypothesis to the first hypothesis. After all, if they didn't become extinct at the end of the cretaceous, the only possible alternative is that they did not become extinct. But science can't work this way: The assertion is not verified - there are no plesiosaur fossils in post-cretaceous deposits, even though it could be verified by the discovery of such fossils. However, it cannot be falsified: there is no evidence which could show that plesiosaurs did *not* become extinct at the end of the cretaceous. We may look for such fossils but never find them because they don't exist, or because we are looking in the wrong places, or because the places in which post-cretaceous plesiosaurs lived did not provide conditions suitable for fossilisation, or for any other number of unknown reasons. This makes is it not a useless hypothesis, but not an hypothesis at all. An inherent element of any hypothesis that it should be falsifiable. Without falsifiability there is no hypothesis. There is an obvious parallel in the discovery of Latimeria. Before 1938, one might have formulated the hypothesis that coelacanths became extinct by the end of the Cretaceous. This hypothesis would be verified by the lack of such fossils, and could be falsified by the discovery of post-cretaceous coelacanths. It was falsified rather dramatically in 1938 when Latimeria was discovered. This does not mean that it was not a good hypothesis - the fact that it was falsified means that it was. Furthermore, it does not mean that we now have to work from the hypothesis that coelacanths survived to the present day: that's a fact, not an hypothesis. The assertion that they have survived to the present day is verified by the existence of Latimeria, but that assertion is not an hypothesis: it cannot be falsified. Latimeria can't be ‘undiscovered’ again.”- Richard Forrest, Something about hypotheses, http://newsgroups.derkeiler.com/Archive/Talk/talk.origins/2006-05/msg05799.html
"Reworked fossils. Fossils from rocks of one age that have been eroded, transported, and redeposited in sediments of younger age."- Geologic Timescale Foundation, https://engineering.purdue.edu/Stratigraphy/strat_guide/bio.html (Editors note: As most reading this will be aware, plesiosaurs were not dinosaurs. However, the current controversy within the paleontological community as to whether Paleocene dinosaur fossils represent dinosaur survival after the Cretaceous mass extinctions or "reworked" Cretaceous fossils is indirectly relevant to the " living plesiosaur" controversy in that there exist many post-Cretaceous "reworked" plesiosaur fossils, spanning from the Paleocene to the Pleistocene.) "Vertebrate fossils have been important for relative dating of terrestrial rocks for decades,but direct dating of these fossils has heretofore been unsuccessful. In this study we employ recent advances in laser ablation in situ U-Pb dating techniques to directly date two dinosaur fossils from the San Juan Basin of northwestern New Mexico and southwestern Colorado, United States. A Cretaceous dinosaur bone collected from just below the Cretaceous-Paleogene interface yielded a U-Pb date of 73.6 ± 0.9 Ma, in excellent agreement with a previously determined 40Ar/39Ar date of 73.04 ± 0.25 Ma for an ash bed near this site. The second dinosaur bone sample from Paleocene strata just above the Cretaceous-Paleogene interface yielded a Paleocene U-Pb date of 64.8 ± 0.9 Ma, consistent with palynologic, paleomagnetic, and fossil-mammal biochronologic data. This first successful direct dating of fossil vertebrate bone provides a new methodology with the potential to directly obtain accurate dates for any vertebrate fossil. A potential weakness in determining the age of a fossil based on the age of the strata containing the fossil is the possibility that the fossil in question may have been reworked from older strata. The direct dating of fossil bone could preclude the reworking hypothesis. We herein report the first successful direct U-Pb age determinations for two fossil dinosaur bones using an in situ laser ablation–multicollector–inductively coupled plasma–mass spectrometer technique(LA-MC-ICP-MS). We believe that the 64.8 ± 0.9 Ma age obtained for the longitudinal section BB1B provides an accurate age for the deposition and diagenesis of this dinosaur bone. Moreover, because this bone was dated directly, it’s age supports the geochemical data indicating that this bone was not reworked from underlying Cretaceous strata. This direct U-Pb age for bone BB1 also provides independent evidence suggesting the possible survival of some dinosaurs into the Paleocene in the San Juan Basin area, as proposed in Fassett (1982, 2009), Fassett et al. (1987, 2002), and Fassett and Lucas (2000)."- James E. Fassett et al., Direct U-Pb dating of Cretaceous and Paleocene dinosaur bones, San Juan Basin, New Mexico, Geology 39(2) (2011), pp. 159-162 "Based on U-Pb dating of two dinosaur bones from the San Juan Basin
of New Mexico (United States), Fassett et al. (2011) claim to provide the first successful direct dating of fossil bones and to establish the presence of Paleocene dinosaurs. Fassett et al. ignore previously published work that directly questions their stratigraphic interpretations (Lucas et al., 2009), and fail to provide sufficient descriptions of instrumental, geochronological, and
statistical treatments of the data to allow evaluation of the potentially complex diagenetic and recrystallization history of bone. These shortcomings lead us to question the validity of the U-Pb dates published by Fassett et al. and their conclusions regarding the existence of Paleocene dinosaurs. Given the importance of accurate and precise age determinations for assigning a Paleocene age to dinosaur bones, we contend that there should have been a more rigorous validation of the dating methods and verification of ages from additional samples beyond those presented by Fassett et al. These weaknesses, combined with the large uncertainty of recrystallization duration of the bones in question, and the contradictory biostratigraphic and magnetostratigraphic placement of the K/T boundary in the San Juan Basin, provide enough doubt to reject the results and interpretations of Fassett et al. We conclude that Fassett et al. have failed to provide the extraordinary evidence needed to support the extraordinary claim that dinosaurs survived the K/T impact event and lived into the Paleocene."- Alan E. Koenig et al., Direct U-Pb dating of Cretaceous and Paleocene dinosaur bones, San Juan Basin New Mexico: COMMENT, Geology 40 (2012), pg. 262 "The finding of what appears to be solid evidence for Paleocene dinosaurs has a number of implications. One implication is that it demonstrates the powerful tendency of scientists to group data into rigid time slots according to the assumed evolution of index fossils. Such tendencies have been labeled the ‘reinforcement syndrome’. The reinforcement syndrome is the tendency by which a hypothesis, or result, is repeatedly reinforced by further data, especially if the hypothesis has been developed by a respected scientist. There are many ways of making data agree with preconceived ideas. Once the idea becomes engrained into science, it is very difficult to dislodge it according to the idea of ruling paradigms developed by Thomas Kuhn. The reinforcement syndrome is a form of circular reasoning, and it is very common in experimental and historical science. The reinforcement syndrome, I believe, is responsible for the belief that dinosaurs finally died out at the end of the Cretaceous, exactly 65 million years ago, and that the mammals took over afterwards. Examples of the reinforcement syndrome are provided when paleontologists simply re-dated ‘Tertiary’ strata to the ‘Cretaceous’ whenever dinosaur remains were found. For instance, dinosaur fossils found in France and India, from what was at first considered Tertiary strata, were subsequently re-dated as Cretaceous. Dinosaurs fossils found in Tertiary strata in eastern Montana have been vigorously opposed and claimed to be caused by reworking."- Michael J. Oard, Paleocene dinosaurs and the reinforcement syndrome, Technical Journal 17 (3) (2003), pp. 5-8 (Editor’s note: A similar controversy to the existence of Paleocene dinosaurs is the controversy over the age and origin of Paleocene elasmobranchs (sharks and rays) found in the Hanna Formation of Wyoming.) "The Hanna Formation, exposed in the northeastern Hanna Basin, Wyoming, represents deposition from late early Paleocene into earliest Eocene time in alluvial, floodplain, and lacustrine environments. A 600-m-thick section that yields abundant vertebrate fossils begins 975 m above the local base of the formation. This section has been dated as latest Torrejonian
through middle Tiffanian using mammalian index fossils. The terrestrial mammals are accompanied by numerous elasmobranch teeth, representing species thought extinct since the end of Cretaceous time. They are species known from two locally widespread marine Cretaceous units, the wall Creek Member of the Frontier Formation and the Steele Shale. These units are broadly exposed flanks on the Sweetwater arch, which define the northern margin of the Hanna Basin. The elasmobranch teeth from the Hanna Formation range in size from a few millimeters to over three centimeters and display transport-induced abrasion not seen in their in situ Cretaceous counterparts. Enameloid of cutting edges, crown points, and cusplets is rounded or sometimes broken, and bony bases commonly are etched or dissolved away. These teeth were reworked and transported from the Wall Creek Member and Steele Shale during uplift and erosion of the Sweetwater arch in middle Paleocene time. Lithic clasts from Paleozoic and Mesozoic strata derived from that arch also occur in association with the elasmobranch teeth in the Hanna Formation. The Cretaceous clasts and teeth indicate a local, northerly source for part of the Hanna Formation in the northeastern Hanna basin."- John H. Burris, Reworked Cretaceous elasmobranch teeth and provenance of the Paleocene Hanna formation (Hanna Basin, Wyoming), Rocky Mountain Geology 36(1) (2001), pp.37-48 "Historically, sandy paleochannel deposits of the 2,000 m-thick Ferris (Maastrichtian-Danian: 66-62 Ma) and overlying 3,000 m-thick Hanna Formation (Danian-Thanetian: 62-55 Ma) in southern Wyoming’s Hanna and Carbon basins have been interpreted as fluvial channel fills. New selachian paleofaunas from both formations are concentrated in stratigraphic intervals that also exhibit an abundance of mechanical tidal indicators and low-diversity marine ichnofossil assemblages (indicating brackish-water). Their presence in these two continental formations supports the reinterpretation of some sandy paleochannel deposits and shaly intervals as brackish-water estuaries, tidally influenced distributary channels, and interdistributary bays. Cederstroemia and Cretorectolobus are reported for the first time from Maastrichtian deposits (they previously have been documented only from Campanian and older strata). A new species of dasyatid is preserved in the middle Ferris Formation (earliest Paleocene: 64 Ma) and awaits fuller description. Prior interpretations of the Hanna Basin area as being far removed from the influences of the Western Interior Sea (WIS) during the latest Cretaceous and Paleocene must now be re-evaluated. New paleogeographic reconstructions of the western shoreline of the WIS, based in part on the stratigraphic and and geographic distribution of selachian teeth in the Hanna Basin area, suggest that a complete marine regression and continental draining at the end of the Cretaceous did not occur. When combined with sedimentological and ichnological data, selachian tooth assemblages are useful tools for interpreting depositional environments and for basin-scale research."- Anton F.-J. Wroblewski, New Selachian Paleofaunas from "Fluvial" Deposits of the Ferris and Lower Hanna Formations (Maastrichtian-Selandian: 66-58 Ma), Southern Wyoming, Palaios 19 (2004),pp. 249-258 "The Arkadelphia Formation-Midway Group contact (Maastrichtian–Paleocene) near Malvern, Arkansas preserves one of the youngest plesiosaur assemblages yet reported from the Gulf Coastal Plain of the United States. The assemblage consists of three vertebrae, a pubis bone and two teeth recovered by scuba diving an outcrop along a meander bend of the
Ouachita River. These plesiosaur remains are preserved in mollusk, coral and ammonoid coquina and may derive from a single individual achieving a total overall length of 10 meters. Taphonomic conditions under which this coquina was deposited indicate that plesiosaurs may have inhabited a shallow, biohermal patch reef environment in southwestern Arkansas where they preyed upon an abundance of ammonoids, bonyfish and chondrichthyans such as Placenticeras sp., Enchodus sp. and Serratolamna serrata. The Arkadelphia Formation-Midway Group plesiosaur assemblage extends the known geographic range of plesiosaurs in North America and indicates that these apex marine reptiles were living at, or near, the K–Pg mass extinction boundary in the region."-Martin A. Becker et al., PLESIOSAUR REMAINS FROM THE ARKADELPHIA FORMATION-MIDWAY GROUP CONTACT (MAASTRICHTIAN-PALEOCENE) HOT SPRING COUNTY, NEAR MALVERN, ARKANSAS, Paper No. 31156-206786 (Abstract Poster),2012 Geological Society of America Annual Meeting and Exposition (Nov.4-7),Charlotte, North Carolina,U.S.A. "Marine reptile elements from the Takatika Grit, Chatham Islands have previously been mentioned but never formally described. This paper describes in detail a new marine reptile assemblage from the Chatham Islands. It is also one of the youngest marine reptile assemblages occurring close to the Cretaceous-Paleogene boundary from the high-latitudes of the Southern Hemisphere. The time slice from the Campanian to the Danian (Late Cretaceous-early Paleogene) witnessed the submergence of a finger-like tract of land informally known as the Chatham Peninsula. The well-constrained New Zealand biostratigraphic record has resulted in a mid-Campanian to mid-Danian age range for the Takatika Grit due to its rich dinoflagellate assemblages, correlated to the zonation proposed for New Zealand by Crampton et al.
(2000). The Takatika Grit comprises mainly mid-Campanian species typical of the Satyrodinium haumuriense Zone plus a slightly less conspicuous Lower Paleocene component (Palaeocystodinium golzowense Zone). This zonation was based on samples that were removed from matrix on and around an unidentified mosasaur from the nodular-phosphorite bone package (NPB).In addition to the Campanian and Danian palynomorphs, Wilson(1982) recorded New Zealand taxa characteristic of the Cretaceous-Paleogene(KPg)boundary. The palynomorphic data indicate that the phosphate nodules and fossils of the NPB are Late Cretaceous in age, but due to sedimentary starvation and reworking, Paleocene microfossil taxa are recorded. Mosasaurine mosasaurs and the elasmosaurid plesiosaurs were all found in the uppermost horizons of the nodular-phosphorite bone package (NPB) along Maunganui Beach, Chatham Island, New Zealand."- Christopher P. Consoli and Jeffrey D. Stilwell, Late Cretaceous marine reptiles (Elasmosauridae and Mosasauridae) of the Chatham Islands, New Zealand, Cretaceous Research 30 (2009),pp.991–999 "McKay (1877c, p. 37) recorded a reptile apparently from Waipara Greensand (Teurian, Paleocene) in North Canterbury, as follows: ‘I was . . . fortunate enough to find portions of the skeleton of a very large Plesiosaurus crassicostatus in a detached mass of green sand ... near the junction of Birchhollow Creek with the Waipara, at the Limestone Gorge. No trace of Saurian remains could be discovered in the higher parts of the green sands . . .".This and other finds apparently in Tertiary sediments of the Waipara area have been discussed widely. Thomson (1920) reviewed discussion of McKay's "Tertiary reptiles", and noted that, in 1913, he also obtained part of a saurian jaw near Birch Hollow, Waipara. Welles & Gregg (1971), who also mentioned these finds, observed that McKay's and Thomson's specimens have not been located, and that they are regarded as having been derived from Haumurian (Late Cretaceous) sediments."- R. Ewan Fordyce, Records of two Paleogene turtles and notes on other Tertiary reptilian remains from New Zealand, New Zealand Journal of Geology and Geophysics 22(6) (1979), pp.737-741



From the above article: "The youngest plesiosaurs with well-established ages are Maastrichtian, but not latest Maastrichtian. Therefore, at present, we take the conservative course, concluding that the Cajon Pass plesiosaurs are of Late Cretaceous age, although there is some evidence, largely unsubstantiated, that one of them could be Paleocene. There is, thus, no conclusive evidence from Cajon Pass that plesiosaurs survived the Cretaceous."- Spencer G. Lucas and Robert E. Reynolds, Putative Paleocene plesiosaurs from Cajon Pass, California, U.S.A., Cretaceous Research 14 (1993), pp.107-111 "STOP A2: COSY DELL FORMATION (K-T BOUNDARY?), Field Presentation by Bob Reynolds. Cajon pass contains 70 million years of history in the rock record. Sediments of Cretaceous age are present as is the Earliest Miocene Vaqueros Fm. Continental deposition starts with the Middle Miocene Cajon Valley Beds. The Inface Bluffs on the northern horizon contain the Brunhes-Matuyama reversal (765,000 yr) suggesting a record of deposition thru the middle-late Pleistocene. At this stop, between I-15 and old Route 66, we are examining the Cretaceous Cosy Dell Formation (Morton and Miller, 2003), formerly called the Paleocene–Eocene? San Franscisquito Formation (Dibblee, 1967; Woodburne and Golz, 1972). The basal conglomerate is overlain by limey sandstones and dark, silty limestone.
It’s Cretaceous age is determined by the presence of elasmosaur vertebrae in the outcrop to the west (Kooser, 1985). Articulated vertebrae in an outcrop to the northwest (Whale Mountain) suggest that this large marine reptile died in place and was not reworked from other sediments (Lucas and Reynolds, 1991). The section may have potential to contain the K/T boundary, but none of the abundant fossil fish scales, crustaceans, gastropods, pelecypods, or plants from this outcrop provide additional age control relating to that transition. Paleocene Coccoplithus pelagicus, a
nanofossil, Apectodinum plexus, a dinoflagellate, and Turritella pachecoensis, a gastropod, were reported (Fred Berry p. c. to Kooser, 1985; Weaver 1951-56) from apparently similar sediments somewhere in Cajon Pass, but by the 1970s the specimens and locality data could not be found."-Geology and Hydrology in the Eastern San Gabriel Mountains: A Journey through the River of Time, Annual Field Trip Number 36, Stops and Discussions, June 19 and 20, 2010
{On the contrary there is no especially good evidence to assume any of the fossils ARE of Cretaceous origin when they are found in Paleocene strata. Their condition is similar to other, presumably contemporary, beach-rolled specimens of Paleocene age and the authors acknowledge the fact. The only reason to assume otherwise is only a blind prejudice that it must be that way]
"A new assemblage of marine vertebrates from northern Saudi Arabia, east of the Nafud, leads us to reconsider the age of the top unit of the Aruma Formation, the Lina Member, hitherto referred to the Maastrichtian. This assemblage contains the remains of a dozen selachian and actinopterygian fishes, as well as those of a giant sea turtle representing a new dermochelyid taxon. It suggests a Late Paleocene to Early Eocene age for this unit. This new dating and a revision of the stratigraphic position of the Lina Member demonstrate the existence, on a regional scale, of an important hiatus at the K/T boundary. This said, it can be underlined that the very poor state of preservation of the ichthyofauna suggests that it is, in part, undoubtably reworked from a Maastrichtian horizon. The existence of reworked material is manifest when seen in the light of the discovery, in the Lina Member, of a rolled ammonite in the form of a cobble along with a single fragment of a plesiosaur tooth. The major lacuna of post-K/T boundary deposits, which may correspond to a major part of the Paleocene, must have reconcentrated, along the Tertiary transgression base, numerous fossils eroded out of the unit lying directly below."- Herbert Thomas et al., Late Paleocene to Early Eocene marine vertebrates from the Uppermost Aruma Formation (northern Saudi Arabia): implications for the K/T transition, C.R. Acad. Sci. Paris. Sciences de la terre et des planetes/Earth and Planetary Sciences 329 (1999), pp. 905-912 "Discosaurus vetustus Leidy- The literature on this species at this point probably outweighs the specimen. The type consists of two battered centra. One, at least, was collected by M. Tuomey and presented to the Academy of Natural Sciences of Philadelphia before 1856. It bears the label "Choctaw Bluff, Clarke County, Alabama", which is certainly in error: this locality is a well-known fossil site but is of Eocene age, millions of years after the extinction of plesiosaurs. Another Choctaw Bluff, in Greene County, is rich in fossil vertebrates and is of Cretaceous age (upper Eutaw and basal Mooreville). It is probably the locality from which this specimen came."- John T. Thurmond and Douglas E. Jones, "Fossil Vertebrates of Alabama" (University of Alabama Press), 1981, pg. 140 (Editors note: The Eocene deposits of Choctaw Bluff, Clarke County, Alabama, are famous regionally for containing the fossils of the Eocene whales Basilosaurus cetoides and Dorudon atrox, see Thurmond and Jones, "Fossil Vertebrates of Alabama".)

Basilosaurus cetoides, also known as Zeuglodon cetoides
Dorudon atrox
"Discosaurus vetustus Leidy……A much mutilated body of a vertebra from Choctaw Bluff, Clarke Co., Alabama, presented by Prof. M. Tuomey of the Academy. The specimen has the same form as just described, excepting that it’s articular faces are more concave and it is considerably larger."-Joseph Leidy, Cretaceous Reptiles of the United States, Smithsonian Contributions to Knowledge 14 (1865), pp.22-25

Discosaurus vetustus vertebrae (Choctaw Bluff specimen unidentified)
"Joseph Leidy had written several articles about the earliest finds of Plesiosaurs in North America, and one of them was the disputed 'Discosaurus' in Alabama, possibly originating from the same beds as 'Zeuglodon' (Basilosaurus). He was writing in the 1850s and one of the comparable early finds was from the Greensands of New Jersey, thought to have been of Late Cretaceous age. The specimens in this case were named 'Cimoliasaurus'; however, some of them turned out to be cetacean vertebrae of probably Pliocene age, probably some
However, this was some of the vertebrae and not all: Leidy did think the other vertebrae were legitimate and were of that genus, and probably related to that.
However, it seems that both genus names are invalid. 'Cimoliasaurus' has been described as a 'garbage taxon' and several nondescript fossils from Europe and Australia have also been ascribed to this genus, much in the same way as the early tendency to call all early canivorous dinosaur finds as Megalosaurus'. http://www.palaeos.com/Vertebrates/Units/220Lepidosauromorpha/220.820.html
In this case the really interesting thing is that the New Jersey fragmentary Plesiosaur is found in association with Pliocene dolphin fossils, mixed up together and only separated out later, and the Alabama fossils Leidy considered probably the same genus are labeled as coming from the Eocene Zeuglodon beds. In the case of the New Jersey Greensands, there is independent evidence that they are not only Cretaceous but also Tertiary: another site gives a paper in which several genera of O. C. Marsh's 'Cretaceous' birds from the New Jersey Greensands are actually of Eocene date or later. http://elibrary.unm.edu/sora/Auk/v084n02/p0260-p0262.html
The characteristics of these fossils has placed them tentatively in the same family as Cryptocleidus and Muraenosaurus, and they were thought to have been like the Elasmosaurs but with shorter necks. This is also along the lines of what the surviving Plesiosaurs would have to have been to give rise to our Long-necked Sea-serpents: long-necked, but not excessively long-necked, not so specialised as the extreme Elasmosaurs, and generalized enough to be versatile, possibly enough so that they could pursue other avenues of evolution that became open to them. That makes a good deal of sense and I am willing to arrange the theory of Plesiosaurian survival on those terms alone."
- Dale Drinnon, On "Discosaurus" and the possibility of Plesiosaurian Survival, Frontiers of Zoology Blog, Jan.8, 2010, www.forteanzoology.blogspot.com/2010/01/dale-drinnon-on–discosaurus-and.html

[At this point Scott Mardis produced several more documents in which Leidy identified the remains of seals and dolphins as Cretaceous inhabitants of New Jersey oweing to their presence in the Greensands levels. These documents although interesting in a general sense do not add anything especially to the case other than confirming that the Greensands layers are indeed geologically ambiguous. Furthermore, Scott has included
several fossils of Plesiosaurs from Glacial deposits, including well-known ones from Germany and Saskatchewan, Canada. Their presence in such deposits are not good indications that Plesiosaurs actually favoured lake habitats along the glacial front, although that is one possible conclusion. I have chosen to leave those examples out. Nor yet do I think it necessary to include documentation that Loch Ness and Lake Champlain were arms of the sea in the Glacial period: this is well known and I do not think it is necessary that Plesiosaurs have g in either lake all of the time since the ice age. DD]
"The Loch Ness plesiosaur: a series of plesiosaur vertebrae has been found on the shore of Loch Ness (July 16, 2003) and hailed by some monster hunters as ‘proof’ of a living plesiosaur in the loch. So, what do we know about this find? Firstly, although it has been confirmed as plesiosaurian by the staff of the National Museums of Scotland (NMS), it contributes nothing to the debate over the existence of a monster in Loch Ness. The bones are millions of years old, and Loch Ness is (in geological terms at least) very recent- only 12,000 years old or so, being a glacially excavated trough. Secondly, it didn’t originate in the loch. As the NMS staff, and also Matt Dale of ‘Mr. Wood’s Fossils’ in Edinburgh, spotted, its general appearance, and the presence of holes made by burrowing sponges, show that it has spent some time in the sea, probably as a beach pebble on a coastal outcrop such as the Jurassic of the Moray Firth coast, or somewhere further afield. Yet Loch Ness is fresh water. I have not had the chance to examine the specimen myself, and have to rely on the photographs published in the popular press (all of which show the specimen upside-down, by the way). It is an articulated series of what look like dorsal or very worn cervical vertebrae. The zygapophyses seem rather low and flat, and the rims of the articular faces rather rounded. The specimen is heavily water-worn, and a lot of surface detail appears to have been lost. If there is any mystery about the specimen, it is how it ended up in Loch Ness. Natural transport as a glacial erratic is unlikely given the distribution of the nearest Jurassic strata, say the NMS staff, who have been diplomatically noncommittal about how it got there. A deliberate hoax has been suggested by several people in the media. There is indeed a long history of hoaxes associated with the Loch Ness ‘monster’ phenomenon, and some perpetrator may well have been willing to waste money probably to no effect. Finds of plesiosaur bones are not very common, and it would take someone who knew something about vertebrate paleontology to recognize them as such at the first finding and, after the specimen was put in Loch Ness, to take the risk that nobody would find it. There is no suggestion that Mr. McSorley was knowingly involved in any such deception. Indeed, the newspaper reports quote him as cleaning off ‘about an inch of green algae’ which suggests it had been lying in the water of the loch for some time. It was, moreover, sheer luck that Mr. McSorley happened to find it when a Dutch tourist, who knew his fossils, was standing by. It would be possible to buy such fossils- I’d guess a specimen of this kind could sell for a couple of hundred pounds. This would make it a rather expensive, and very shallow, deception. However, a credible alternative is that the fossil was used as a demo piece for tour groups to show what real plesiosaur bones look like, and was left behind by accident one day. This actually happened before, in the case of a plesiosaurian limb bone in the 1980’s. It will be interesting to watch the story develop."- Richard Forrest, The Loch Ness Plesiosaur, The Plesiosaur Site, www.plesiosaur.com/plesiosaurs/lochnessplesiosaur.php


Gerald McSorley and Loch Ness plesiosaur vertebrae
" The fossil clearly shows four perfectly preserved vertebrae, complete with spinal chord, and blood vessels, set in grey limestone. …Mr. McSorley, 67, found the fossil after stumbling in shallow water near the bank of the loch. ‘I literally tripped over the fossil in the water,’ he said. ‘When I put my hands down to steady myself I saw something unusual and picked it up. Once I had cleaned off about an inch of green algae, and I could see the texture of the bone, it became clear I had an important fossil.’ - Pensioner finds ‘Nessie ‘ Fossil, BBC News, July 16, 2003 " ‘The fossil is embedded in a gray, Jurassic-aged limestone. Rocks in the Loch Ness area are much older- they’re all crystalline, igneous and metamorphic rocks’.
(Lyall) Anderson (NMS) says the nearest match is at Eathie on the Black Isle, some 30 miles (50 kilometers) northeast of Loch Ness".- Loch Ness Sea Monster Fossil a Hoax, Say Scientists, National Geographic News, July 29, 2003
(Editor’s note: Though limestone is usually associated in people’s mind with the Mesozoic era, late Quaternary limestone deposits are known. The sand deposits in Fort Augustus from late Quaternary flooding, in combination with carbonates from the shells of marine invertebrates from Loch Ness’s interglacial and post-glacial marine phases, could theoretically have produced a sandstone or limestone matrix around a Quaternary-aged fossil.) "The 260 m ice-dammed lake in glens Spean and Roy finally attained a volume of 5 km3. It is suggested that it was initially drained subglacially and catastrophically through the Spean gorge and Loch Ness. Maximal flow of water may have been about 22,500m3s-'. In Glen Spean the ice dam, 7 km long and up to 200 m high, collapsed. Near Fort Augustus a vast deposit of sand and gravel was laid down in relation to a Loch Ness suddenly raised in level.
Subsequently, the level of the Spean-Roy lake varied constantly as it was intermittently
emptied by other sudden floods of lesser volume, some of which followed the Lundy gorge and one of which produced a now abandoned waterfall site near Loch Lochy. Some of these events are related to the formation of end moraines and fluvial terraces in the area around Spean Bridge and Gairlochy."- J. B. Sissons, Catastrophic lake drainage in Glen Spean and the Great Glen, Scotland, Journal of the Geological Society of London 136(1979), pp. 215-224 "As a setting for the reporting of our additional findings, we first call attention to the earlier-mentioned evidence of others that the previous Great Ice Age of Europe had receded over a hundred thousand years ago and left a long interglacial period until the development of the most recent glacier. This may be highly significant to our further disclosure that, by amino acid reaction-rate dating, Boomer et al. (2002) have reported,2 and verified under different temperature hypotheses, that the ancient sea-bed-matrix material we recovered appears, indeed, also to be coincidentally in the range of about 125,000 years old! In trying to reconcile the order-of-magnitude difference in this dating from the 14C dating, we observe that while the amino acid results are known to be beyond the range of 14C dating, is it conceivable that the younger dating could result from
some phenomenon such as the re-precipitation of shell carbonates?"- Robert H. Rines and Frank M. Dougherty, Proof Positive—Loch Ness Was an Ancient Arm of the Sea, Journal of Scientific Exploration 17(2) (2003), pp. 317–323 "The diagenetic history of the pre-LGM (Last Glacial Maximum) limestone from the core by IODP Expedition 310 was investigated texturally and geochemically. The limestone is mostly deposited in reef front and fore-reef settings and mostly composed of corals, coralline algae, and microbialites with a minor contribution of green algae, mollusks, and echinoderm fragments. Relatively high abundance of microbialites is the quite distinctive feature, compared to other reefal sediments described elsewhere. Texturally, shallow marine cements are characterized by acicular and botryoidal aragonite and druse HMC, and equant LMC cements of meteoric origin are relatively rare. Presence of LMC equant cements appears to be facies-controlled, thus is dependant upon the nature of substrates. It appears that the limestone was mostly subjected to vadose meteoric diagenesis during the last glacial period. The fibrous HMC cements in vuggy pores together with their oxygen isotopic compositions and textural evidence suggest that the limestone was subaerially exposed during the last glacial period and resubmerged during deglaciation. Stable isotopic and trace elemental analyses were carried out for microbialites that were originally composed of HMC. Oxygen isotopic compositions and Fe and Sr contents of pre-LGM microbialites are clearly distinguished from those of post-LGM limestone. This suggests that the limestone had undergone meteoric diagenesis in the different diagenetic system, depending upon the magnitude of the partition coefficients of trace elements. No clear difference between unaltered (post-LGM) and altered (pre-LGM) limestone was observed for Mn, Mg, and carbon isotope, indicating a closed diagenetic system. However, the diagenetic system was semi-open with respect to Sr and oxygen isotope. In addition, enriched carbon isotopic compositions of the pre-LGM limestone may imply that vegetation cover was negligible for the formation of paleosol layers. This is also supported by poorly preserved unconformity surface with an absence of paleosol layers within the core."- Kyung Sik Woo et al., Carbonate diagenesis of the Late Pleistocene limestone of the Faaa M0020A core: IODP Expedition 310, Tahiti Sea Level Change, Geosciences Journal 14(2) (2010),pp. 225-234
"The ‘molluscs in muddy sand with occasional corals’ particularly Manicina areolata, represent the Thalassia community that is still common to the Belize lagoons (e.g. Macintyre et al., 2000). Thus although this Pleistocene limestone has been greatly altered and much of the original texture lost in our cores, it appears that the Holocene mangroves of Twin Cays became established on a Pleistocene substrate formed by lagoonal accumulations of branching Porites moundsand Thalassia beds, when this substrate was flooded by the Holocene Transgression."-
IAN G. MACINTYRE and MARGUERITE A. TOSCANO, THE PLEISTOCENE LIMESTONE FOUNDATION BELOW TWIN CAYS, BELIZE, CENTRAL AMERICA, ATOLL RESEARCH BULLETIN NO. 511, ISSUED BY NATIONAL MUSEUM OF NATURAL HISTORY SMITHSONIAN INSTITUTION
WASHINGTON, D.C., U.S.A. SEPTEMBER 2004
(Editor’s note: If sea urchins existed in Loch Ness during it’s marine period, I see no reason why burrowing sponges could not have been there, as well. Sponges were certainly in the Champlain Sea, a close analogue to a marine Loch Ness)
............
[Here Scott Mardis included several of his comparison charts including photos of the Loch Ness Monster and The Lake Champlain Monster, and skeletons of Plesiosaurs. Since these were already covered separately, I did not think it was necessary to rerun them here-DD]
(Editor’s conclusion: Make of this what you will. It is apparent to all that scientific acceptance of the reality of any strange animals existing in places like Loch Ness or Lake Champlain is contingent on one criteria: the acquisition of unambiguous biological evidence for such an animal. Entertaining the idea of the possibility of the reality of such animals by no means entails embracing the "relict plesiosaur" theory, as many clever alternatives exist. Having said that, it is not unreasonable to say that, sifting through the alleged evidence from Loch Ness and Lake Champlain, a significant number of people have claimed to have seen animals or objects that vaguely resemble long-necked plesiosaurs and that there also exists photographic evidence associated with these bodies of water of objects that also vaguely resemble long-necked plesiosaurs. The "relict plesiosaur" theory has been endorsed in the past by several prominent scientists including Robert Rines, Denys Tucker and Sir Peter Scott. It has generally been ridiculed in the paleontological community for obvious reasons: the discovery of a post-Mesozoic or living plesiosaur would be a great embarrassment. The excitement of such an astounding discovery would probably be far outweighed by the negative impact on orthodox geologic theory. I suspect that if unambiguous evidence of a living plesiosaur were discovered tomorrow, the very same people who ridicule the idea with impunity now would back peddle and instantly create a new structure of theory to account for it. But this is only speculation. Relative to Loch Ness in particular, the anti-plesiosaur argument goes like this: "There is no geologic evidence for the survival of plesiosaurs, which were marine animals, after 65 million years ago and Loch Ness, a fresh water lake, has only existed since the end of the last glacial cycle, 12,000 years ago. Before that, it was filled with glacial ice and nothing could have been living in it." On the surface this sounds all cut and dried, but the true reality is more nuanced. Loch Ness and Lake Champlain both sit inside deep, ancient geologic faults (380 Mya and 440 Mya, respectively) and the odds are very high that there were large bodies of water in both places for eons before the glacial cycles. Cambrian-Ordovician period marine invertebrate fossils from 500 to 440 million years old are abundant on islands in and along the shorelines of Lake Champlain. When the glacial cycles did fill Loch Ness and Lake Champlain with glacial ice, aquatic animals living in them died, were driven into the sea or to so-called glacial refugia, bodies of fresh water beyond the glacial margins. There is evidence, particularly from North American fishes and freshwater invertebrates, that these animals reinvaded their former ranges during glacial flooding in the interglacial periods. This seems to have happened multiple times. The marine invertebrate fossils discovered in Loch Ness and two sets of 14C and aminoacid racemization ages for them (12.8 Kya and 125 Kya) seem to suggest that Loch Ness was subject to two separate marine incursions between glaciations. The multiple whale and pinniped fossils from Champlain Sea deposits attest to the fact that what would become Lake Champlain was open to the sea as recently as 10,000 years ago. The presence of rainbow smelt, landlocked Atlantic salmon and sea lampreys in modern Lake Champlain is proof that some animals survived the transition from the Champlain Sea to Lake Champlain. Some of the invertebrates in Loch Ness are glacial relicts (the bivalve Pisidium conventus and the chironomid Sergentia coracina). As it has reasonably been established that both Loch Ness and Lake Champlain were subject to invasion by marine animals in the geologic past, the antiquity of animal lineages in the open sea was not constrained by the presence of glacial ice and marine animals with ancient, pre-glacial pedigrees could have made their way into these bodies of water. The most famous of ancient survivors is the Coelacanth, believed extinct for 80 million years until rediscovered alive in 1938. Some fossil coelacanths were freshwater fishes and the living representatives retain osmoregulatory features that could theoretically facilitate survival in freshwater. The Comoro Islands where Latimeria chalumnae survives today are only 5 million years old. This implies that the ancestors of the Comoro Island coelacanths wandered the seas for some 75 million years before the volcanic birth of the Comoros. What if they had wandered to the coast of Scotland instead? What if the first living coelacanth had been caught in Loch Ness? Looking at the litany of "reworked" plesiosaur fossils spanning from the Paleocene to the Pleistocene, I think the "no geologic evidence of plesiosaurs after the Cretaceous" argument is shot to hell. True, there is no unambiguous post-Cretaceous evidence that is not fragmentary. Nevertheless, there is some evidence. If only one isolated tooth, flipper bone or vertebra of a plesiosaur is found in a Mesozoic deposit, the paleontologists do not immediately invoke reworking to account for it’s presence in the strata where it was found. But let the same bone be found in a Cenozoic deposit, complete with other Cenozoic marine vertebrates, and it is immediately tossed into a refuse bucket labeled "reworked’. Instead of marveling over this
potential relict survivor, it is barely mentioned in the literature with disdain and then shoved into a drawer, hidden away to be forgotten or conveniently lost. If "reworking" will not get the job done, then one can always say it was mislabeled or even misidentified. In the case of the Cajon Pass plesiosaurs, what began to look like promising evidence for Paleocene plesiosaurs was swept under the rug by simply magically turning the Paleocene San Francisquito Formation into the Cretaceous Cozy Dell Formation. There is a term for this. It’s called "moving the goal posts". No doubt that the arch-nemesis of the Great Sea Serpent, Sir Richard Owen, would hardily approve. Speaking of sea serpents, the long history of sightings of vaguely plesiosaur-like animals at sea (sometimes by scientists such as Meade-Waldo and Nicoll in 1905) should provide an answer for the potential marine ancestors of these theoretical long-necked "lake monsters". I’ve no doubt that reworking of fossils is a very real phenomenon. The mechanisms are usually tidal and fluvial erosion, tectonic activity, glacial scouring, erosion by wind and rain, volcanism, burrowing by animals on land and in the sea, even dislodgement by tree roots. But one begins to get the impression that this is a convenient and easy out whenever a problematic fossil is found. Geology is a very conservative science. Many once-controversial ideas that are now generally accepted in geology were once vigorously opposed: plate tectonics, the Bretz Floods, the descent of birds from dinosaurs, the idea that dinosaurs were warm-blooded, a bolide impact cause for the K/T boundary extinctions. The small minority of paleontologists that question whether "reworked" Paleocene dinosaurs are in fact not reworked but genuine post-Cretaceous survivors are being vigorously opposed at the present time. If this minority is eventually proven right, perhaps the day will come when these anomalous "reworked" plesiosaur fossils will be reexamined in a different light. It is probably too much to hope for the paleontology community to bear these anomalous fossils in mind before ridiculing the slight possibility that people may be really encountering living descendants of the plesiosaurs in these "monster" sightings. The negative argument against the potential reality of sea serpents/lake monsters is short and to the point: to date, no biological evidence for such animals can be examined in a laboratory anywhere in the world. There exists peripheral circumstantial evidence, including lost potential biological evidence, that some, including myself, find compelling. Brilliant people, like Robert Rines, can believe in the possible reality of improbable unorthodoxies that may eventually be proven wrong and still be brilliant. The courage to explore unpopular ideas without fear of failure has sometimes led to astounding discoveries. The current state of research into the natural history of plesiosaurs has gone a long way toward dispensing with the "water’s too cold for plesiosaurs" and "plesiosaurs didn’t live in freshwater" arguments. Other theoretical negative arguments, largely independent of the plesiosaur theory, such as "not enough food", "not enough genetic diversity through inbreeding", "would be seen at surface breathing often", "dead bodies would be conspicuous" and "no place to hide from sonar" remain just that, theoretical. If these animals exist, we really know nothing of their natural history. Can we really be sure we know that much about the ancient plesiosaurs, much less their hypothetical descendants 65 million years removed or possibly some totally different group of animals that may only coincidentally resemble plesiosaurs?
The consistent failure of ROV’s to intercept potential sonar contacts in time to identify them visually has led me to the conclusion that the only effective answer is minisubmarines or AUV’s. You’ve got to go down to where these animals are living most of the time if you want to find them. And most expeditions to these lakes are usually for only a few days at most. The odds are low that positive evidence is going to appear in such a short window. Time and the proper equipment are probably going to cost a lot of money. Until the day that someone is willing to bankroll such a long shot, little progress is likely to be made. In this context, it is not so ludicrous to ponder why so little progress has been made in 80 years at Loch Ness. Still, one of the most enthusiastic partisans of the Loch Ness "Monsters", Robert Rines, methodically searched the bottom of Loch Ness and found no mortal remains of what is popularly assumed to be an isolated breeding colony of animals that have been inhabiting Loch Ness for 10.000 years. Except possibly this.































{kind=link}